Важни фактори на еволюцията според биолога Сергей Сергеевич Четвериков

От 1926г.
Установено е, че гените не са абсолютно стабилни. Редица причини водят до промяна в структурата им, местоположението в хромозомите и самия хромозомен набор.
Голяма част от измененията водят до възникването на нежизнеспособни форми, които загиват на определен стадий от своето развитие.
Друга част поставят "бариера", която възпрепятства кръстосването.
Има и такива, които се включват в генния фонд и се предават от поколение на поколение. При процес на мейоза те претърпяват всички препятствия на генетичната рекомбинация и в определен момент се проявяват във фенотипа.
Още преди години се е стигнало до заключението, че видът постоянно се обогатява с мутантни алели. По-труден е бил за обяснение фактът, че видовете остават стабилни в продължение на много поколения. Както е известно, по-голямата част от мутациите са рецесивни. Всяка мутация се приема за рядко срещано явление.
След размножаването си в следващите поколения, шансът да срещне два еднакви гена в зиготата е по-голям. Смятало се е, че процесът може да бъде ускорен от селекционера - практически всички породи домашни животни и сортове културни растения са хомозиготни по различни рецесивни гени.
При кръстосването на два сорта от тях много често в потомството се получавало връщане към изходния "див" тип. Това е било известно на Ч. Дарвин след кръстосването му на различни породи гълъби и получаването на тъмносив цвят със синкав оттенък. В този случай механизмът е бил идентичен със случая със супресорните мутации - дефектът на един геном се оправя от друг.

Биологът Четвериков е стигнал до извода, че най-важно значение за еволюцията имат само тези изменения на генетичния материал, които, намирайки се в рецесива, не се проявяват във фенотипа. Без да се подлагат на действието на отбора, те могат да се натрупат в популацията в достатъчно голямо количество.
Изключение тук били бактериите и синьозелените водорасли - хаплоидни организми с неоформено ядро, при които липсва мейоза. Притежават само един набор от гени и затова всяка мутация се проявява назабавно във фенотипа, нямат запас от генетична изменчивост.
Приспособявайки се, те не еволюират, а остават такива каквито са били, когато са били единствените обитатели на планетата. Поради тази причина вирусите и бактериите са изработили свои механизми на генетична рекомбинация, които били далеч от мейозиса и половия процес при висшите организми. Установено е, че не всяка мутация, която е възникнала наново, е можела да се утвърди в потомството.
Съществувал и обратния процес, обедняване или загуба на генетичния фонд. Вероятността за загуба на мутацията е можела да се пресметне - ако се приеме, че от потомството на една самка оживяват два индивида, т. е. числеността на популацията не се увеличава и не се намалява (това, което немците наричат система на две деца - Zweinkin-der system), то съществуват 37 шанса на сто, че тази мутация ще се загуби.
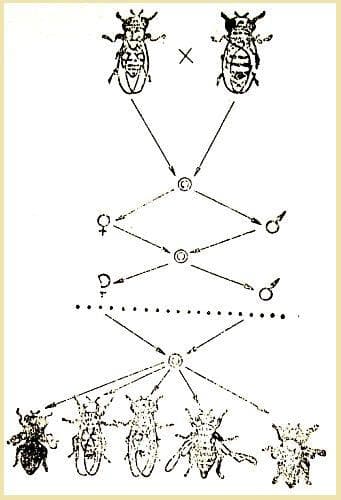
Опит на С. Четвериков - в потомството на една дрозофила след редица поколения при близкородствени кръстоски се отцепват рецесивни мутанти хомозиготи, които дотогава са се намирали в скрит вид в еднообразният т.нар. "див тип".
Заедно с мутационния процес, като важен фактор на еволюцията, биологът Сергей Четвериков е отличил числените колебания в една популация, които образно нарекъл "вълни на живота".
Като пример за това е посочен фактът, че през ранната пролет всеки от нас вероятно е виждал презимувала муха между рамките на своя прозорец, докато през месец август те са изключително активни.
Също така има години, когато нашествията от комари например са често срещано явление и години, когато тяхната численост е много малка. Голямата численост на някои видове понякога има планетарен характер. Такива са например масовите миграции на скакалците в годините на усиленото им размножаване, годините на "мише нашествие" и др.

Част от тези явления са циклични: имат сезонен, годишен или многогодишен период. Други са апериодични. Това са явления на нарастване числеността на организмите, попаднали в нова среда на обитаване с малък брой врагове, грипните епидемии и редица други заболявания, бързо разпространяващите се нашествия на вредители от други страни по Земята.
Като друг пример може да се посочи колорадският бръмбар, чиято родина е Северна Америка. Там, използвайки за храна диви растения от семейство картофови, той не може да достигне голяма численост на своята популация. Славата си на вредител е завоювал на европейските картофени полета. Същото се отнася и за филоксерата (американската лозова въшка).

Невинаги резките подеми в числеността на видовете са се приемали за вредни. Например бурното развитие на азовския червей "нереис" в Каспийско море е увеличило значително "фуражната" база на рибите. Това от своя страна е компенсирало обедняването на Каспийско море в резултат на урегулирането на стичането на р. Волга. Установено е, че ако е нямало "нереис", отслабването на Северно Каспийско море е щяло да нанесе много вреди на рибите там.
Интересни факти могат да се отбележат за колебанията в числеността, когато период на максимум е милион пъти по-голям от периодите на минимум. Смятало се е, че "вълните на живота" могат да променят популационния генофонд, изменяйки числеността на всички алели. Това след себе си би довело до промяна в свойствата на популацията.
Установено е, че при намаляване на "вълните на живота", числеността на определени популации се увеличава, а следователно нараства и вероятността за среща на рецесивите в хомозиготите; скритите алели се проявяват във фенотиповете и попадат под действието на естествения отбор.
Новата "вълна на живота" би се състоялa от индивиди с изменен генофонд. При спадане на "вълната" част от мутациите просто отпадат от генофонда. Ефектът е наречен "гърло на бутилка", т.е. не всички са можели да преминат в новите поколения чрез минимална численост. Така например, ако от една популация лисици изчезнат индивидите, които са носители на гените за черно-кафява окраска, става невъзможна появата на черни лисици в тази популация.

Установено е също, че при спадане на "вълната на живота" дребните популации, изолирани в миналото, ще бъдат обединени в една и ще започне активна генна обмяна. Във връзка с това Четвериков е описал много интересен феномен, който теоритикът Е. Майр е нарекъл "принцип на основателя".
Нови ареали (например един отдалечен остров) могат да бъдат заселени от случайно докарани там индивиди от нов за ареала вид. Пример за това е западноевропейската популация от ондатри (мускусен плъх), водеща началото си от пет индивида, които са пуснати в началото на 20 век от Прага.

В заключение се е стигнало до извода, че "принципът на основателя" води до възникването на нови видове, подвидове и раси.
Като важен фактор, съпътстващ еволюцията и оказал влияние в нейното развитие биолога Сергей Четвериков е посочил и ролята на изолацията, ограничението на панмиксията*, което произтича по различни причини.
Една от тези причини се дължи на факта, че при понижаване "вълната на живота" (също един от важните фактори на еволюцията според Четвериков) настъпва разпадане на популацията на изолирани една от друга субпопулации.
Получените "парчета" са наречени "изолати" и тяхната изолация е била още по-голяма при наличието на физически прегради. Като пример може да се посочат човекоподобните маймуни. Тъй като те не могат да плуват (днес учените са установили точно обратното), за тях дори и най-плиткото поточе би било едно много трудно и непреодолимо препятствие. Значението на пространствената изолация с времето е започнало да се надценява.

Вероятността за кръстосване в популацията се ограничава от други форми на изолацията. Една от тези форми е екологичната изолация. При нея се намалява възможността за среща на индивиди, готови за размножаване. Като пример могат да се посочат сезонните раси растения, риби и голяма част от безгръбначните, при което изолацията не настъпва в пространство, а във време.
Като пример за съвместно съществуване в една и съща популация на форми, които се размножават едновременно, но не се кръстосват помежду си, може да се посочи дрозофилата. При нея има "жълта мутация". Сред сивите самки, жълтият самец има много малки шансове за успех, защото вибрира с крилата си много бавно, а това от своя страна намалява неговата привлекателност.
Ако тези крила му бъдат отрязани, той напълно ще изгуби своята способност да привлича самките и да се чифтосва с тях. Напротив, дрозофолни самки с отстранени мустачки-антени са по-малко "претенциозни" и се чифтосват със самци от други видове.
Има описани редица подобни случаи за най-различни животни. Дори ако чифтосването (опрашването при растенията) е осъществено, обикновено оплождане не може да се получи. Прието е тази форма на изолация да се означава като "физиологична". Има редица растения, чийто прашец не може да прорасне върху рилцето на пестика на едни растения, но за сметка на това прораства на други.
Третата и може би най-важна форма на изолация е "генетичната". В този случай оплождане може да се осъществи, но хибридите от първо поколение няма да бъдат жизнеспособни (или стерилни) поради нарушаване на мейозата.
Смятало се е, че тези типове на изолация могат да бъдат преодоляни чрез различни методи (например чрез изкуствено осеменяване). Издигането на бариера от генетичната изолация между формите се е смятало за граница на вида, което грешно. Редица видове не са се смесвали с близки видове само благодарение на различия в брачното поведение. Може да се опише и обратният процес - лесно е да бъде изведена форма, която да е генетично изолирана от изходната.
Установено е, че изолацията закрепва различията в малките популации (субпопулации) и без нея е невъзможно съществуването на дивергенция - при промяна, ще се измени цялата популация и един вид би преминал в друг, без възникването на нови. Това не е видообразуване, а видопревръщане.
Дълго време е отнело на науката да разгадае точния смисъл и значение на изолацията - дали е причина или резултат на генетични различия. Отговорът разбира се няма как да бъде еднозначен. Като пример могат да се посочат зайците, аклиматизирани на на о. Мадейра, при които е настъпила толкова голяма промяна, че са загубили способността си за кръстосване с европейските. В този случай генетичните различия се приемат като "страничен ефект" от натрупването им за времето на разделното съществуване на популациите.
Съществуват много други случаи на малки генетични различия, които водят до изолация, която пък разцепва популацията толкова, че се стига до формирането на видове. Като пример за генетични различия може да се посочи промяната в честотата на брачно квакане при жабите, а също и промяната в скоростта на блещукане при светулките.

Четвериков е отстоявал своята теза, че изолацията не създава нови видове, а се развива постепенно и еволюцията е резултат от действието на група фактори.
***
*панмиксия - кръстосване на индивиди с различни генотипове в рамките на популацията. Прието е, че панмиксията не е абсолютна в природата.
*дивергенция - раздалечаване на популационните генофондове в резултат на явления като естествен отбор, генетичен дрейф и мутиране.
Продукти свързани със СТАТИЯТА

ПРОИЗХОД НА ВИДОВЕТЕ - САБИНА РАДЕВА - СИЕЛА

НАЙ-ВЕЛИКОТО ШОУ НА ЗЕМЯТА - РИЧАРД ДОКИНС - СИЕЛА
Библиография
"Еволюцията днес", Б.Медников
СТАТИЯТА е свързана към
- Етапи от историята на човешката еволюция
- Хомо сапиенс - единственият оцелял човешки вид
- История на биологичната еволюция
- Луси - най-известният австралопитек на света
- История на човешките раси
- Хомо еректус - първият, напуснал пределите на люлката на човечеството Африка
- Какво е "Луси"? Бързи факти за нашия прародител - австралопитека
- Хомо хабилис вероятно е първият наш предшественик, изработвал каменни инструменти
- Ардипитекус рамидус като част от еволюцията на човека
- Полинезийските островитяни като част от историята на човешката еволюция
Коментари към Важни фактори на еволюцията според биолога Сергей Сергеевич Четвериков